

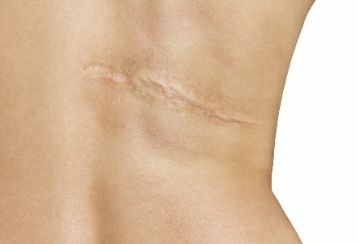
二、典型无瘢痕创面愈合
人类的胎儿时期存在无瘢痕愈合的功能(基于该特性、唇腭裂患儿经官腔镜修复术后无明显瘢痕存留),自然界 中,同样存在着具有皮肤创面无瘢痕再生愈合的生物,这种自然存在的生物学功能可能存在于该生物的整个生命周期 或其中 1 个特定时期,如斑马鱼、部分两栖动物和非洲多刺小鼠,这些动物在整个生命周期中皮肤创面均具有无瘢痕再生愈合的功能。通过对这些无瘢痕创面愈合过程的共性的研究,可能获得对人类创面和瘢痕治疗方式的突破性进展。
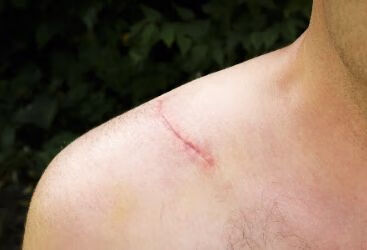
蛋白酶和胶原酶重塑,持续的胶原沉积和分解代谢慢慢取代 肉芽组织,但沉积在创面床内的胶原蛋白不会恢复到受伤前 状态【16-17]。在个体正常发育期间,胶原蛋白被加工成网状 编织图案使组织中具有弹性,机体损伤后的胶原蛋白以线性 方式沉积失去原始的网状编织图案及弹性118-20]。在重塑过 程结束时,瘢痕的拉伸强度仅达到完整皮肤的70%,因此导
致坚韧、僵硬的瘢痕组织
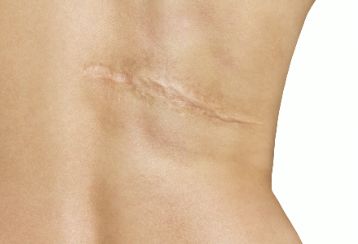
三)瘢痕形成
成纤维细胞是在创面床内产生 ECM 以重建真皮的主要细胞类型。成纤维细胞产生的第 1 个主要 ECM 成分是纤连蛋白,但成纤维细胞在创面中的增殖和迁移减慢后,蛋白质的产生就转向 I 型和 III 型胶原蛋白,而至少在受伤 3 个月后才会产生沉积弹性蛋白纤维。此外,成纤维细胞在转化生长因子-p(transforming growth factor-p,TGF—p )的作用下分化成肌成纤维细胞并产生大量的 ECM。肌成纤维细胞与胶原纤维结合并导致创面收缩。
胶原蛋白作为瘢痕组织最主要的 ECM 成分,在创面床中被成纤维细胞、嗜中性粒细胞和巨噬细胞分泌的基质金属
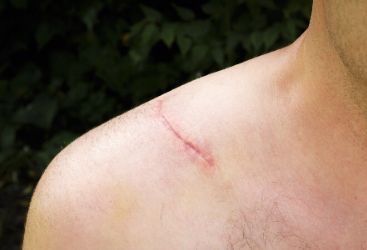
阶段中细胞外基质(extracellular matrix,ECM )的组成成分及含量,如胶原蛋白、透明质酸、弹性蛋白等都会明显发生变化。至今的研究尚未完全探明皮肤发育的每个阶段中成纤维细胞的基因表达与转录变化,成纤维细胞和 ECM 成熟的分子机制方面仍无突破性的研究进展,具体机制和变化过程仍处于不明确状态。
(二)皮肤受损与修复
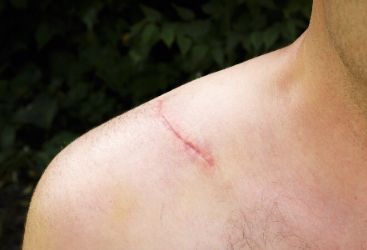
当皮肤受到物理损伤时,皮肤的稳态转换暂时停止,转而进行创面愈合和修复反应。创面的严重程度将决定最终愈合结果,简单的划痕可以有效修复,但全层皮肤创面常常导致瘢痕形成。然而,当人体的皮肤发生了严重损伤后,不能从严重损伤的病理改变恢复其原始功能,而是形成修复性瘢痕。人类和大多数哺乳动物的创面愈合由 3 个重叠阶段组成:炎症,增殖和成熟/重塑,最终导致纤维化反应。
皮肤表皮由角质形成细胞(其构成细胞表皮的 80%一 90%,形成对抗外部环境的屏障)、黑素细胞(合成黑色素,保护皮肤不受紫外线伤害)、朗格汉斯细胞(免疫细胞,防止感染)和默克尔细胞(参与触觉功能)构成 Ho。表皮层内通过肿瘤蛋白 p63 阳性表皮祖细胞在基底层产生 1 个 p63 阳性子细胞维持增殖;在基底上的分化层产生 1 个 p63 失表达子细胞,并终末分化为提供结构和保护的角质化框架,达到表皮层的稳态。5.0o。
皮肤真皮是经过 4 个阶段发育形成的结缔组织层,每个
着生活、生产、交通,以及放化疗、慢性疾病、人口老龄化的发展,皮肤作为人体最外层的物理防御屏障最先遭受伤害,而皮肤创面往往最终以瘢痕的形式愈合。2 J。大自然中存在着皮肤无瘢痕愈合的机制,本文将围绕皮肤无瘢痕愈合的研究进展进行综述。
一、皮肤创面愈合与瘢痕形成
(一)皮肤稳态
皮肤是人体最大的器官,起着多种重要作用,包括调节温度、预防感染等。皮肤是一种高度动态的器官,整个生命过程处于相对活跃的状态。皮肤主要由外胚层分化形成的表皮层和由中胚层分化形成的真皮层组成。
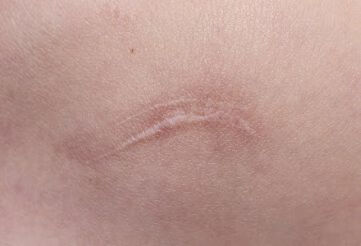
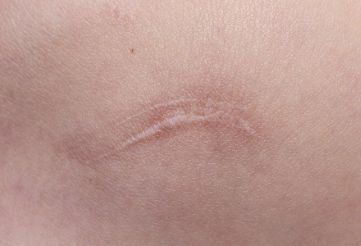
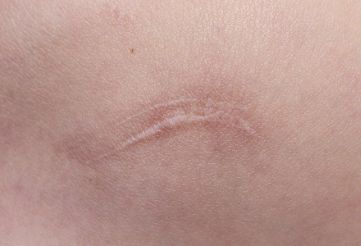
5 结论 通常认为最为理想的组织修复方法是再生式,而瘢痕愈合代表着组织再生失败。瘢痕愈合与组织再生均可观察到,例如将成体哺乳动物的肝脏组织切除 2/3 后,剩余的 1/3 肝脏组织会完全再生,但是肝脏穿刺后的伤口组织会呈现瘢痕组织愈合【24】。使用微小的针刺伤皮肤表面或者大面积纹身均是以组织修复方式愈合,但是小面积切割伤会以瘢痕愈合方式修复,并且于其他部位相比,胸骨、胸肩部位出现的瘢痕程度更深。由此可见,再生与瘢痕愈合 2 种方式均存在于成体之中,因损伤方式、损伤部位、组织类型等方面存在的差异,导致瘢痕严重程度、修复方式也不尽相同。因此,目前医者面临的问题是如何抑制瘢痕愈合基因位点表达,诱导皮肤组织再生基因位点表达,促进胶原的有序排列,达到无瘢痕愈合。
大分子透明质酸在年龄增长的影响下,价值透明质酸酶和活性氧的作用,就会变成不同大小的片段。高分子透明质酸在伤口愈合期间对炎性反应和纤维组织增生有很好的抑制作用,降低纤维化产生。而低分子透明质酸及其残余片段,则可引发纤维化,产生炎性反应,破坏相应组织,影响伤口愈合。
透明质酸激活 P13 激酶/AKT 和 MEKl/ERKl—2 是通过介导细胞表面 RHAMM/HMM,CD44,LYVE l 和 ToU 样受体(TLR )2/4 受体激活,从而对纤维化产生进行抑制。若受体出现表达异常,将影响伤口愈合。诸如伤口愈合早期 ERKl—2 表达缺失,可延缓成纤维细胞、间充质细胞迁移分化速度,延迟炎性反应及血管化,减慢伤口愈合速度;CD44 受体异常表达,可降低胶质细胞增殖速率,降低炎性反应,减慢上皮化速率;TLR2/4 受体缺如可延迟伤口愈合【12。14J。
交联降低,继而改善瘢痕出现。有研究指出,胎鼠 在孕龄17 d与孕龄19 d时赖氨酰氧化酶的含量不 同,其中17 d时只有一点点增加,而在19 d时增加 2倍且保持很久(四)。临床抑制产生瘢痕是通过铜离 子与D一青霉胺、B一氨基丙腈螯合,从而达到对赖氨酰氧化酶抑制的目的。细胞外基质中透明质酸是重要部分。透明质酸 是一种糖胺聚糖,有葡糖醛酸和葡糖双糖结构反复 交联而成,主要参与细胞的分裂、分化、炎性细胞的 游走、迁移及细胞因子合成等,在伤口愈合中,能够 起到抗炎、调控胶原合成、促进伤口愈合的作用
肽链上的醛基(一 CHO )是胶原蛋白分子内与分子间形成共价键的桥梁,从而相交连接组成胶原纤维。
胶原酶喜欢破坏没有组成交联的多肽链分子结构。胶原合成中的限速酶为脯氨酰羟化酶。成纤维细胞在不同阶段分泌脯氨酰羟化酶的量不同,成人和胚胎晚期的分泌量显著低于胚胎早期。
在 20 孕周以前,其含量比较高,后逐渐下降,至出生后与成人水平一致,促进胶原交联的是赖氨酰氧化酶【9l。对赖氨酰氧化酶的活性进行抑制,从而使得胶原蛋白
展开更多



